Анатомія
Розглядаючи біомеханічні принципи функціонування кульшового суглоба людини, необхідно враховувати нормальну анатомію проксимального відділу стегнової кістки і таза, оскільки м’язи, зв’язки, а також кісткові структури забезпечують рівновагу сил, що дозволяє здійснювати контрольовані рухи в стегново-ацетабулярному з’єднанні. Хоча локальна анатомічна структура кульшового суглоба має велике значення, необхідно зазначити, що кульшовий суглоб з’єднує тулуб і нижню кінцівку, і ці структури суттєво впливають на біомеханіку кульшового суглоба.
За класичним визначенням, кульшовий суглоб — це обмежене з’єднання між кулястою голівкою проксимального відділу стегнової кістки і вертлюжною западиною тазової кістки. Разом голівка стегнової кістки та вертлюжна западина утворюють кулястий суглоб. Вертлюжна западина розміщується в місці з’єднання клубової, сідничної та лонної кісток таза — Y-подібний хрящ — і починає розвиватися в ембріона на 8-му тижні [1], а на 11-му тижні кульшовий суглоб вже є повноцінно сформованим [2]. Зростання хрящової пластини відбувається через Y-подібний хрящ, при цьому ямка кульшової западини розвивається донизу, убік та вперед у відповідь на наявність головки стегнової кістки [3, 4]. Розміри та орієнтація ацетабулярного отвору описуються рентгенографічно: оцінюються латеральний центральний кут, передній центральний кут, кут Tonnis, а також кут ацетабулярного нахилу; значення цих кутів залежить від глибини вертлюжної западини, нахилу її поверхні, що утримує вагу, а також відповідного обсягу покриття головки стегнової кістки [5]. Розвиток вертлюжної западини продовжується до зростання Y-подібного хряща, зазвичай до віку від 16 до 18 років [3].
Хоча класичне визначення кульшового суглоба як кулеподібного суглоба з єдиним центром ротації є концептуально простим уявленням і, таким чином, дозволяє робити припущення щодо біомеханічних властивостей суглоба та робити робочі підрахунки, треба пам’ятати, що голівка стегнової кістки має не ідеально сферичну форму. Сам центр ротації не однаковий для різних видів руху. Крім того, індивідуальні відхилення в морфології проксимального відділу стегнової кістки впливають на ступінь того, наскільки це явище може бути спостережене, і дозволяють зробити припущення щодо можливості розвитку коксартрозу в майбутньому [6]. Незначні відмінності в морфології стегнової кістки та морфології вертлюжної западини впливають на біомеханіку кульшового суглоба, але ці відмінності ще не були описані докладно, що ускладнює розрахунки, саме тому в цій статті ми виходимо з того, що кульшовий суглоб є єдиним кулеподібним суглобом з одним центром ротації та сферичною головкою стегнової кістки, якщо інше не встановлено.
Функціональна глибина вертлюжної западини збільшується за рахунок вертлюжної губи, що складається з фіброволокнистого хряща, що є краєм вертлюжної западини і знаходиться глибоко в суглобовій сумці. Така структура підвищує функціональну стійкість суглоба, діючи як ущільнювач між западиною та голівкою стегнової кістки, запобігаючи зникненню синовіальної рідини зсередини суглоба [7, 8]. Щоб від’єднати голівку стегнової кістки від вертлюжної западини, треба подолати дію ущільнювача, що додає кульшовому суглобу стійкості [9]. Це призводить до того, що в останках тіла людини, коли зв’язки та м’язи стегна розщеплюються, стегнова кістка може залишатися в западині через ефект засмоктування [10]. Крім того, види діяльності, пов’язані з навантаженнями, за наявності діючого ущільнювача вертлюжної губи призводять до збільшення внутрішньосуглобового тиску — під час цього процесу внутрішньосуглобове тертя зменшується за рахунок покращання змащування суглоба [7, 9].
Куляста голівка стегнової кістки розвивається одночасно з вертлюжною западиною [3] і пов’язана з діафізом стегнової кістки через її внутрішньокапсульну шийку та позакапсульні міжвертельні ділянки. Нормальний нахил між шийкою стегнової кістки та її тілом (шийково-діафізарний кут) поступово зменшується від народження до моменту, коли скелет є вже повністю сформованим. У дитини, яка щойно народилася, цей кут становить 150 градусів, а в повністю сформованому скелеті дорослої людини — 125 ± 5 градусів [11]. В аксіальній площині шийка стегнової кістки має ротацію допереду (антеверсія) щодо дистальної осі надвиростків стегнової кістки, причому нормальним вважається антеверсія від 12 до 14 градусів [12]. Оскільки ступінь антеверсії проксимальної частини стегнової кістки збільшується, також зростає механічна перевага великого сідничного м’яза [13], при цьому механічні можливості привідних м’язів зменшуються [14]. Великий і малий вертлюги — це виступи на відповідно крайньому боковому і задньосередньому проксимальному відділі стегнової кістки, де з’єднуються різні м’язи, які відповідають за рух стегна.
Кульшовий суглоб оточений товстою волокнинною капсулою, що починається з вертлюжної губи в бік передньої міжвертлюгової лінії проксимального відділу стегнової кістки спереду і до задньої міжвертлюгової лінії ззаду [11]. Із капсулою кульшового суглоба пов’язані три зв’язки, які допомагають стабілізувати його в екстремальних точках руху. Серед цих зв’язок: іліофеморальна зв’язка (зв’язка Bigelow), яка простягається від передньої нижньої клубової ості до передньої міжвертлюгової лінії, лобково-стегнова зв’язка, яка приєднується до верхньої лобкової частини та нижньої поверхні шийки стегнової кістки, а також сіднично-стегнова зв’язка, яка простягається від сідничної кістки до задньонижньої поверхні шийки стегнової кістки. Ці три зв’язки стабілізують голівку, відповідно, під час гіпернапруження, гіперабдукції і розтягування. При цьому іліофеморальна зв’язка найміцніша з них, а задня сіднично-стегнова зв’язка найтонша і найслабша [14]. Ці зв’язки щільніші, коли кульшовий суглоб знаходиться в розігнутому положенні, що пояснює збільшення амплітуди обертання під час згинання порівняно з розгинанням.
Постачання крові до проксимального відділу стегнової кістки переважно здійснюється кінцевими гілками присередніх огинаючих артерій стегна, спереду — у меншому обсязі через бічну огинаючу артерію [15]. Вертлюжна западина і вертлюжна губа кровопостачаються з передньої сідничної, нижньої сідничної та затульної артерій [11, 14, 16, 17].
Під час рухів у кульшовому суглобі задіяна дуже велика кількість м’язів, що також відповідають за правильне розташування нижніх кінцівок та тулуба відносно один одного. Тип руху, на який впливають такі м’язи, залежить від його походження, кріплення м’язів та положення кульшового суглоба стосовно таза та коліна.
Клубово-поперековий м’яз, прямий м’яз стегна, кравецький м’яз і м’яз-натягувач широкої фасції відповідають за згинання стегна в кульшовому суглобі. Перші три іннервуються стегновим нервом, а м’яз-натягувач широкої фасції — верхнім сідничним нервом [11].
Розгинання стегна в кульшовому суглобі здійснюється шляхом скорочення великого сідничного м’яза і м’язів задньої поверхні стегна (біцепс стегна, півперетинчастий та півсухожилковий м’язи) [11]. Великий сідничний м’яз іннервує нижній сідничний нерв, а м’язи задньої поверхні стегна — великогомілкова гілка сідничного нерва [11]. Скорочення середнього сідничного м’яза, малого сідничного м’яза та м’яза-натягувача широкої фасції іннервуються гілками великого сідничного нерва та призводять як до скорочення, так і до зовнішнього обертання голівки стегнової кістки у вертлюжній западині. Аддукція стегна в кульшовому суглобі досягається скороченням великого привідного м’яза, довгого привідного м’яза, короткого привідного м’яза, які іннервуються затульним нервом. М’язи, задіяні у внутрішньому обертанні кульшового суглоба, та відповідні нерви, що їх іннервують, такі: внутрішній затульний м’яз (внутрішній затульний нерв), зовнішній затульний м’яз (затульний нерв), верхній близнюковий м’яз (внутрішній затульний нерв), нижній близнюковий м’яз (нерв квадратного м’яза стегна), грушоподібний м’яз (нерв грушоподібного м’яза S1 та S2), а також квадратний м’яз стегна (нерв квадратного м’яза стегна) [11].
Рухи в кульшовому суглобі
У сагітальній площині в нормальному кульшовому суглобі може здійснювати згинальний рух на 120–125 градусів, а розгинальний рух — на 10–15 градусів [12, 18]. Загалом такий рух обмежується капсульно-зв’язковими структурами і м’язово-сухожильними елементами, а також кістковою архітектурою стегна. Згинання стегна значною мірою обмежено положенням колінного суглоба, тоді як розгинання колінного суглоба значно знижує згинання стегна через пов’язане зменшення напруження м’язів задньої поверхні стегна, які проходять повз обидва згадані суглоби [12]. Ілеофеморальна зв’язка, передня капсула і м’язи-згиначі обмежують розгинання кульшового суглоба [12]. Коли стегно знаходиться в положенні згинання від 0 до 70 градусів, бічне обертання може бути від 0 до 90 градусів [19]. Коли в кульшовому суглобі стегно знаходиться в положенні розгинання, присереднє (внутрішнє) та бічне (зовнішнє) обертання значно менші, оскільки м’які тканини стегна знаходяться в стані більшого напруження під час розгинання, таким чином обмежуючи ступінь обертання.
Поєднаний рух кульшового суглоба і таза разом впливає на весь рух стегна, а амплітуда рухів, згадана вище, залежить також і від руху таза. Дюберрі та співавтори описують відмінності в задньому обертанні таза: з колінним суглобом у положенні згинання 26% згинання стегна відбувається в результаті попереково-тазового обертання, а з колінним суглобом у положенні розгинання у результаті попереково-тазового обертання відбувається 39% згинання стегна [18]. Вважається, що під час піднімання ваги вплив обертання таза на згинання стегна становить 18 % [20].
Хоча основні обмеження амплітуди руху кульшового суглоба традиційно пов’язують із м’якими тканинами стегна, останнім часом значний інтерес становить роль кісткової архітектури. За умов, які призводять до аномалій проксимальної стегнової або ацетабулярної морфології, наприклад фемороацетабулярний імпінджмент, рух часто обмежений через аномальний контакт кісток між ділянками проксимальної частини стегнової кістки та вертлюжної губи в крайніх положеннях амплітуди руху [6]. У таких клінічних ситуаціях рух, що відбувається між стегновою кісткою та вертлюжною западиною, є меншим, а таз починає рухатися раніше.
Всі рухи стегна, необхідні для виконання стандартної щоденної діяльності, описані в статті [21] та наводяться в табл. 1.
Джонсон і Смідт провели електрогонометричні дослідження 33 пацієнтів та виявили, що для здійснення щоденної діяльності, наприклад, для зав’язування шнурків, необхідне відведення та бічне (зовнішнє) обертання на 20 градусів, а також згинання стегна на 120 градусів [21, 22]. Крім того, із збільшенням віку пацієнтів відбувається загальне зниження амплітуди руху стегна, при цьому в старших пацієнтів спостерігаються знижена здатність до згинання та розгинання стегна, а також зменшення довжини кроку [20].
Серед деяких категорій атлетів спостерігається зниження амплітуди руху стегна. Наприклад, деякі бігуни на довгі дистанції, які не належать до спортивної еліти і виявляють гнучкість нижчу, ніж в інших атлетів, показують вищий рівень ефективності бігу. Вважається, що вища економічність руху таких спортсменів пов’язана з відносною відсутністю витрат енергії на стабілізацію стегна і таза м’язами та сухожилками під час бігу порівняно з більш гнучкими бігунами [23, 24]. Також розгинання стегна в професійних хокеїстів на 10 градусів менше за норму для їх віку [25], і деякі вчені вказують на скорочення згинання стегна як на потенційну причину хронічного болю в нижньому відділі спини в деяких спортсменів [27].
Цикл ходіння
Кульшовий суглоб поєднує нижні кінцівки та тулуб, і рух, який здійснюється цими частинами тіла, дозволяє ходити на двох ногах. Рух, що здійснюється в кульшовому суглобі під час циклу ходіння, характеризується згідно з різними фазами циклів бігу та ходіння, причому середнє значення згинання стегна та його розгинання відповідно 35 та 10 градусів [22, 28]. Під час ходіння кульшовий суглоб максимально розгинається під час відриву п’яти від землі і максимально згинається наприкінці фази переносу [29]. Під час фази опори в ходінні спостерігається приведення стегна в кульшовому суглобі, а також його присереднє (внутрішнє) обертання, центр гравітації при цьому знаходиться ближче до кульшового суглоба, а під час фази переносу спостерігається відведення та бічне (зовнішнє) обертання, коли нижня кінцівка підтягується допереду. У табл. 2 наводяться зміни положення стегна, задіяних м’язів, їх участь у циклі ходіння та на різних етапах ходьби [30].
/105-1.jpg)
Наявність етапу подвійної опори визначає діяльність ходіння. Коли швидкість ходіння зростає, зменшується тривалість фази подвійної підтримки, а біг починається, коли ця фаза виходить із циклу ходьби і замінюється фазою польоту, коли обидві ноги одночасно відриваються від землі [31]. Із збільшенням швидкості бігу тривалість фази польоту зростає. Ступінь згинання і розгинання стегна змінюється, коли збільшується швидкість ходіння від безпосередньо ходіння до легкого бігу та бігу, причому максимальне згинання стегна збільшується більше ніж до 55 градусів посередині фази переносу стегна під час ходіння та легкого бігу [30]. Під час ходіння та легкого бігу розгинання стегна менше, ніж під час бігу, і відрив пальців ноги від землі відбувається перед максимальним розгинанням стегна під час бігу та легкого бігу, хоча під час ходіння це відбувається після максимального розгинання стегна [30]. Ступінь відведення та приведення кульшового суглоба варіює, коли відбувається прискорення руху від ходіння до легкого бігу та бігу, причому максимальні значення цих рухів спостерігаються під час бігу [30]. Приведення стегна в кульшовому суглобі на максимальній швидкості бігу становить від 15 до 20 градусів і спостерігається відразу перед опусканням п’яти, а максимальне відведення відбувається у фазі переносу під час бігу після відриву пальців ноги від землі [30].
Сили, що впливають на кульшовий суглоб
Впродовж багатьох років багато зусиль спрямовується на описання різних сил, що впливають на реакцію кульшового суглоба людини. Існують оцінки на основі підрахунку та схем діючих сил, але такі методи вимагають припущень та спрощень щодо м’язів-антагоністів та багатьох м’яких тканин, які впливають на розподіл сил у кульшових суглобах. Пряме вимірювання цих сил до сьогодні залишається практично неможливим. Проте прямі вимірювання були здійснені після ендопротезування кульшового суглоба, і ці дані можна застосовувати до нативного суглоба людини.
Розрахунки in vitro
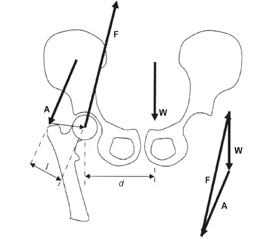
Підрахунки сил, які діють на кульшовий суглоб, є типовими в літературі з біомеханіки та ортопедії, при цьому більшість схем є розкладом сил, що діють поперек кульшового суглоба під час стояння на одній кінцівці (рис. 1) [19, 32, 33]. Такі підрахунки є спрощенням, що грунтується виключно на сили у фронтальній площині, які діють на кульшовий суглоб у статичних умовах, але вони корисні як легка для розуміння основа для демонстрації того, як незначні зміни в положенні тіла або в анатомії стегна впливають на сили, які діють на кульшовий суглоб. У статичних умовах для утримання положення таза паралельно до землі на таз та кульшовий суглоб діють такі сили (рис. 1): сила гравітації W, що дорівнює різниці маси тіла та маси протилежної нижньої кінцівки; А, що є силою відвідних м’язів, під дією яких утримується таз, і F, що є силою, прикладеною головкою стегнової кістки до вертлюжної западини, або силою реакції суглоба [32]. Силу реакції суглоба F можна визначити за силою відведення. Якщо відома маса тіла людини, плече сили гравітації d та плече сили м’язів відведення l, то силу відведення A можна вирахувати за рівнянням (1) [19]:
/106-1.jpg)
У положенні рівноваги сума векторів сили A, F та W дорівнює нулю, отож при додаванні векторів A та W величина сили реакції суглоба F, за підрахунками, у 2,7 раза більша від маси тіла і спрямована під кутом 69 градусів до горизонтальної лінії при стоянні на одній нозі, з паралельним до землі положенням таза [19].
Хода Тренделенбурга спостерігається, коли пацієнт нахиляється у фронтальній площині під час фази стояння чи ходіння таким чином, що більша частина його ваги знаходиться над ногою, на якій він стоїть. Це часто можна спостерігати в пацієнтів із болем у стегні і часто вважається показником основної патології кульшового суглоба або слабкістю відвідного м’яза. Хоча чиста слабкість відвідних м’язів і є нетиповою клінічною знахідкою (за відсутності основної патології кульшового суглоба), розуміння основ біомеханіки кульшового суглоба допомагає пояснити, чому така техніка ходіння дає переваги при болі в стегні. Коли вага людини переміщується на опорну ногу і ближче до центру обертання суглоба, плече сили гравітації скорочується, таким чином знижуючи силу, що має генерувати м’яз відведення, щоб протидіяти силі гравітації, яка діє на таз. Такий результат загального зниження сили/навантаження на кульшовий суглоб при його скороченні пропорційно ступеню скорочення плеча сили гравітації [32]. Іншими словами, чим гірше нахил Тренделенбурга, тим нижчий рівень навантаження на кульшовий суглоб.
Оскільки зміни в значенні плеча сили гравітації співвідносяться з моделлю ходи Тренделенбурга, вони можуть змінюватися шляхом застосування палиці для зменшення навантаження на кульшовий суглоб. Якщо палицю тримають у протилежній руці під час ходіння, генерується сила, направлена догори, що допомагає протидіяти силі гравітації, яка діє на вагу пацієнта, таким чином зменшується сила відведення, необхідна для утримання рівня таза, і відповідно зменшується навантаження на кульшовий суглоб [32]. Значення сили реакції палиці на землі, необхідної для зменшення сили реакції протилежного кульшового суглоба, пропорційно набагато менше через те, що плече сили палиці значно довше за плече сили протилежних відвідних м’язів. Таке зменшення сили можна вирахувати, і деякі дослідники визначають його як приблизно 20 % [32]. Іншими авторами було встановлено, що, максимально збільшуючи ефект використання палиці, можна досягти до 42 % зменшення м’язової активності, що відповідає зменшенню сили реакції кульшового суглоба з 3,4 раза більше маси тіла до 2,2 раза [34].
Хоча наявність і лікування остеоартроза кульшового суглоба не є основним завданням спортивної травматології, у роботах, присвячених їй, постійно згадується, що зменшення маси може значно скоротити силу реакції кульшового суглоба, таким чином потенційно знизити симптоми пацієнтів з артрозом кульшового суглоба. На рис. 1 показано, як на силу реакції кульшового суглоба впливає маса тіла пацієнта і як будь-яке зменшення маси тіла може допомогти знизити цю силу. Використання палиці може бути прикладом для демонстрації пацієнтам користі зменшення маси; полегшення від використання палиці еквівалентно зниженню маси тіла приблизно на 20 % [32].
/106-2.jpg)
Через те, що сила реакції кульшового суглоба пропорційна силі, що прикладена до проксимальної стегнової кістки м’язами відведення (рівняння (1)), ключовим фактором, який впливає на значення сили реакції суглоба, прикладеної до головки стегнової кістки, є відношення плеча сили відвідного м’яза до плеча сили гравітації або відношення l до d у рівнянні, наведеному вище [19]. Коли це відношення зменшується, сила реакції суглоба збільшується. Значення плеча сили гравітації і відвідних м’язів використовуються в реконструктивній хірургії кульшового суглоба. Наприклад, надаючи серединне положення вертлюжному компоненту під час тотального ендопротезування кульшового суглоба, плече сили гравітації може знизити відвідну силу на 40 % і відвідний момент на 50 % [35]. Аналогічно підвищення компенсації стегнової кістки або розворот великого вертлюга боком під час тотального ендопротезування кульшового суглоба підвищує механічну перевагу відвідних м’язів шляхом збільшення плеча сили відвідних м’язів, що співвідноситься із зниженням реакції сили стегна [36]. Остеотомія при варусних деформаціях справляє такий самий ефект, знижуючи силу реакції суглоба шляхом збільшення плеча сили відвідних м’язів.
Хоча результати вивчення сил, що діють на кульшовий суглоб у статичному положенні, корисні та досить чіткі, дія руху та різних видів навантаження, найімовірніше, є більш цікавою та доречною для лікування спортсменів. Сили, що діють на кульшовий суглоб, можна розраховувати для бігу та ходіння, але такі розрахунки потребують врахування кінетичних та кінематичних даних всієї нижньої кінцівки, є досить складними і не є метою цієї роботи. Брікманн та ін. [32] наводять підсумок власної роботи [37], а також роботи Новачека [38], Вінтера [39], Пола [40, 41] і Моррісона [42] щодо таких розрахунків. Ці дослідники показали, що під час повільного ходіння навантаження на кульшовий суглоб досягає близько 3-кратної маси тіла людини відразу після контакту п’яти із землею, і це навантаження збільшується до 4-кратної маси тіла в момент перед відривом пальців ніг від землі [28]. При збільшенні швидкості ходіння пікове значення навантаження на кульшовий суглоб збільшується приблизно до 4 мас тіла людини, а навантаження в момент перед відривом пальців ноги збільшується не так значно.
Під час бігу сили, що дорівнюють від 7 до 8 мас тіла, проходять через кульшовий суглоб під час контакту п’яти із землею і трохи більше — під час відриву пальців ніг від землі [28]. Необхідно зазначити, що ці розрахунки описують рух по прямій, не враховуючи види діяльності, які виконують інші спортсмени, наприклад, у баскетболі, тенісі, футболі, які вимагають виконання різкої зміни напрямку руху.
Ускладнені рухи, такі як підрізання та обертання, характерні для деяких видів спорту, роблять проблематичним розрахунок сили, але ван дер Богерт та співавтори застосували математичну модель, щоб приблизно розрахувати сили, прикладені до кульшового суглоба під час спуску на лижах. Вони виявили, що модуль навантаження на кульшовий суглоб становить від 8,3 до 12,4 маси тіла людини, а за приблизними розрахунками навантажень під час інших видів лижного спорту ці показники менші (від 4,1 до 7,8 маси тіла людини) [43]. Як і очікувалося, біг на лижах створює набагато менше навантаження на кульшовий суглоб, а саме від 4,0 до 4,6 маси тіла людини [43].
Макнітт-Грей зі співавторами, провівши заміри сили реакції опори під час приземлення з 3 різних висот, виявили, що сили реакції опори під час приземлення є нижчими в тренованих гімнастів порівняно зі спортсменами-аматорами. Вони також виявили, що треновані гімнасти досягали найвищого показника сили реакції опори в середньому на 6,3 мс швидше, при цьому аматори пристосовувалися до приземлення з більшої висоти за рахунок збільшення кута згинання кульшового суглоба і загальної тривалості фази приземлення, а професійні спортсмени показували більший момент розгинання гомілково-ступневого суглоба та кульшового суглоба під час приземлення [44].
Вимірювання in vivo
Прямі заміри сил, що діють на кульшовий суглоб спортсменів, потребують встановлення вимірювача сили хірургічним шляхом, що є неможливим. Але Бергман та співавтори зробили спробу описати сили, що діють на ендопротез кульшового суглоба, провівши заміри під час різних видів діяльності після тотального ендопротезування кульшового суглоба [37]. Цій групі дослідників вдалося встановити датчики тиску в компоненти ендопротеза кульшового суглоба. Результати цих вимірювань підтвердили in vitro розрахунки. Як очікувалося, сила, що діє на кульшовий суглоб, під час прискорення ходьби становить: до 300 % ваги людини під час повільного ходіння, 350–400 % під час швидкого ходіння, до 500 % під час легкого бігу і максимально — 800 % — під час спотикання. Цікаво відзначити, що значення сили, що діє на кульшовий суглоб під час стояння на двох ногах, становить від 80 до 100 % ваги людини. Той факт, що ці сили не становлять менше половини цього значення, пов’язують із безперервним напруженням м’язів, що діє на кульшовий суглоб під час такого типу стояння [32]. Перевага такого типу вимірювання — це те, що враховуються всі сили, які діють на кульшовий суглоб, на відміну від in vitro розрахунків, коли дія м’язів або зовсім не враховується, або враховується лише мінімально, а також не враховується пружний натяг м’язів, сухожилків та суглобової капсули [32].
Тиск на суглобові поверхні
Через те, що зміна місця тиску на суглобовий хрящ кульшового суглоба вважається фактором, що впливає на розвиток остеоартрозу кульшового суглоба, корисно розглянути анатомічні елементи і сили, які визначають ці потенційні зміни. Основний тиск на суглоб можна розрахувати як відношення сили реакції суглоба до площі поверхні головки стегнової кістки, на яку діє така сила:
де А — «проектна площа» суглоба. Хоча при цьому не враховується нерівна поверхня головки стегнової кістки і формула є занадто спрощеною, однак, якщо маса тіла становить 60 кг, а діаметр головки стегнової кістки — 5 см, то, використовуючи інформацію зі схем діючих сил, що наведені вище, при силі реакції суглоба 1500 N середнє значення тиску на кульшовий суглоб становить, згідно з розрахунками, 5 N/см2 [32].
Проте реальна робоча поверхня кульшового суглоба не є ідеально сферичною, і реальний розподіл тиску по суглобовому хрящу нерівномірний. Це пов’язане з тим, що кульшова западина має бічний отвір і не є цільною півкулею. Коли ми маємо цільну півкулю, максимальний тиск на суглоб прикладений до точки, що лежить на векторі, який паралельний до вектора сили реакції стегна. Від цієї точки тиск зменшується в напрямку периферії півкулі і досягає нульового значення на губі півкульної западини, якщо в системі немає тертя (що є слушним припущенням, враховуючи, що коефіцієнт тертя кульшового хряща дуже незначний). У такій ситуації, коли немає повного покриття кулі та западини суглоба, за умов рівноваги спостерігається більший тиск на хрящ біля непокритої ділянки губи, а на більш обмеженому боці значення тиску наближені до нуля [32]. Грінвальд та Брінкманн у подальшому описали розподіл тиску як косинусоподібний і змогли розрахувати значення розподілу тиску в півкулях при різних ступенях покриття [45, 46]. Наприклад, у разі кулі з покриттям у 130 градусів точка максимального тиску спостерігається збоку за умов рівноваги, а при зменшенні значення покриття до 110 градусів (симуляція дисплазії кульшового суглоба) максимальний тиск спостерігається на латеральній частині губи, де це значення є більшим, ніж при покритті у 130 градусів [32, 45]. У клінічному відношенні ці розрахунки добре відповідають спостереженим випадкам посилення зносу хряща в пацієнтів із дисплазією суглоба, коли спостерігаються надмірне навантаження на край западини і передчасна поява артрозу. Були зроблені прямі заміри тиску на суглоб у суглобових поверхнях, але виникли певні труднощі. Рушфельт та співавтори вимірювали суглобовий тиск за допомогою ультразвукового зонда на поверхні імпланта-хряща після проведення геміартропластики на зразках кульшового суглоба. Через невеликі розбіжності між діаметром протеза та западини виявилось неможливим зробити загальні висновки щодо замірів тиску природного суглоба людини [47]. Браун і Шоу використовували датчики тиску, встановлені на головку стегнової кістки трупа для вимірювання контактного суглобного тиску, але виявили, що максимальний тиск розподілений випадково [48]. Брінкман описав декілька недоліків в їх експериментах, а саме головка стегнової кістки перебувала під навантаженням у напрямку вектора навантаження і не рухається вільно; спостерігалась невідповідність між жорсткістю матеріалу датчика та суглобового хряща, а також субхондральної кістки, що могло вплинути на результати замірів при навантаженні на систему [32].
Висновки
Базовий опис біомеханіки стегна не задовольняє вимогам спортивної травматології і потребує врахування комплексної взаємодії кісткової структури, м’язів, капсули та зв’язок цього суглоба. Для виявлення та лікування в спортсменів аномалій кульшового суглоба, що спостерігаються в м’яких тканинах стегна, необхідно враховувати кісткові аномалії, бо останнім часом все більше дослідників визнають їх впливовим фактором у встановленні діагнозу та лікуванні хвороб кульшового суглоба.
Конфлікт інтересів. Автори заявляють про відсутність конфлікту інтересів при підготовці даної статті.
Список литературы
1. Rao J., Zhou X.Y., Villar R.N. Injury to the ligamentum teres: mechanism, findings, and results of treatment // Clin. Sports Med. — 2001. — 20. — Р. 791-799.
2. Watanabe R.S. Embryology of the human hip // Clin. Orthop. — 1974. — 98. — Р. 8.
3. Ponseti I.V. Growth and development of the acetabulum in the normal child // Bone Joint Surg. Am. — 1978. — 60. — Р. 575-585.
4. Weinstein S.L. Developmental hip dysplasia and dislocation // Morrissy R.T., Weinstein S.L., eds. Lovell & Winters’ Pediatric Orthopaedics. — 6th ed. — Philadelphia: Lippincott Williams & Wilkins, 2006. — Р. 987-1033.
5. Clohisy J.C., Carlisle J.C., Beaule P.E. et al. A systematic approach to the plain radiographic evaluation of the young adult hip // JBJS Am. — 2008. — 90. — Р. 47-66.
6. Ganz R., Parvizi J., Beck M. et al. Femoroacetabular impingement. A cause for osteoarthritis of the hip // Clin. Orthoped. — 2003. — 417. — Р. 112-120.
7. Ferguson S.J., Bryant J.T., Ganz R. et al. The influence of the acetabular labrum on hip joint cartilage consolidation: a poroelastic finite element model // J. Biomech. — 2000. — 33. — Р. 953-960.
8. Takechi H., Nagashima H., Ito S. Intra-articular pressure of the hip joint outside and inside the limbus // Nippon Seikeigeka Gakkai Zasshi. — 1982. — 56. — Р. 529-536.
9. Johnson J.D., Noble P.C., Hurwitz D.E. et al. Biomechanics of the hip // Callaghan J., Rosenberg A., Rubash H., eds. The Adult Hip. — 2nd ed. — Philadelphia: Lippincott Williams & Wilkins, 2007. — Р. 81-90.
10. Kapandji I.A. The Physiology of the Joints, v. 2. — Edinburgh: Churchill Livingstone, 1970.
11. Robbins C.E. Anatomy and biomechanics // Fagerson T.L., ed. The Hip Handbook. — Boston, MA: Butterworth-Heinemann, 1998. — Р. 1-37.
12. Hamill J., Knutzen K. Biomechanical Basis of Human Movement. — 3rd ed. — Baltimore: Lippincott Williams & Wilkins, 2009. — Р. 187-254.
13. Radin E.L. Biomechanics of the human hip // Clin. Orthop. — 1980. — 152. — Р. 28-34.
14. Sim F.H., Rock M.G., Scott S.G. Pelvis and hip injuries in athletes: Anatomy and function // Nicholas J.A., Hershman E.B., eds. The Lower Extremity & Spine in Sports Med. — 2nd ed. — St Louis, MO: Mosby, 1995. — Р. 1025-1065.
15. Trueta J., Harrison M.H.M. The normal vascular ana–tomy of the femoral head in adult man // J. Bone Joint Surg. Br. — 1953. — 35. — Р. 442-461.
16. Hansen A. Anatomy and surgical approaches // Morrey B., ed. Reconstructive Surgery of the Joints. — 2nd ed. — New York: Churchill Livingsone, 1996. — Р. 889-890.
17. Seldes R.M., Tan V., Hunt J. et al. Anatomy, histologic features, and vascularity of the adult acetabular labrum // Clin. Orthop. — 2001. — 382. — Р. 232-240.
18. Dewberry M.J., Bohannon R.W., Tiberio D. et al. Pelvic and femoral contributions to bilateral hip flexion by subjects suspended from a bar // Clin. Biomech. — 2003. — 18. — Р. 494-499.
19. Nordin M., Frankel V.H. Basic Biomechanics of the Musculoskeletal System. — 3rd ed. — Baltimore: Lippincott Williams & Wilkins, 2001. — Р. 203-221.
20. Murry R., Bohannon R., Tiberio D. et al. Pelvifemoral rhythm during unilateral hip flexion in standing // Clin. Biomech. — 2002. — 17. — Р. 147-151.
21. Johnston R.C., Smidt G.L. Hip motion measurements for selected activities of daily living // Clin. Orthop. — 1970. — 72. — Р. 205.
22. Johnston R.C., Smidt G.L. Measurement of hip-joint motion during walking. Evaluation of an electogoniometric me–thod // J. Bone Joint Surg. — 1969. — 51A. — Р. 1083.
23. Craib M.W., Mitchell V.A., Fields K.B. et al. The association between flexibility and running economy and sub-elite male distance runners // Med. Sei. Sports Exerc. — 1996. — 28. — Р. 737-743.
24. Gleim G.W., Stachenfeld N.S., Nicholas J.A. The influence of flexibility on the economy of walking and jogging // J. Orthop. Res. — 1990. — 8. — Р. 814-823.
25. Tyler T., Zook L., Brittis D. et al. А new pelvic tilt detection device: roentgenographic validation and application to assessment of hip motion in professional ice hockey players // J. Orthop. Sports Phys. Ther. — 1996. — 24. — Р. 303-308.
26. Offierski C.M., Macnab M.B. Hip-spine syndrome // Spine. — 1983. — 8. — Р. 316-321.
27. Ingber R.S. Iliopsoas myofascial dysfunction: a treatable cause of “failed” low back syndrome // Arch. Phys. Med. Rehabil. — 1989. — 70. — Р. 382-386.
28. Kabada M.P., Ramakaishnan H.K., Wooten M.E. et al. Repeatablilty of kinematic, kinetic and electomygraphic data in normal adult gait // J. Orthop. Res. — 1989. — 7. — Р. 849-860.
29. Murray M.P., Kory R.C., Clarkson B.H. Walking patterns in healthy old men // J. Gerontol. — 1969. — 24. — Р. 169-178.
30. Hughes P.E., Hsu J.C., Matava M.J. Hip anatomy and biomechanics in the athlete // Sports Med. Arthrose Rev. — 2002. — 10. — Р. 103-114.
31. Mann R.A. Biomechanics of running // Mack R.P., ed. American Academy of Orthopedic Surgeons Symposium on the Foot and Leg in Running Sports. — St Louis, MO: Mosby, 1982. — Р. 1-29.
32. Brinckmann P., Frobin W., Leivseth G. Musculoske-letal Biomechanics. — New York: Thieme New York, 2002. — Р. 69-84.
33. Pauwels F. Biomechanics of the Locomotor Apparatus: Contributions on the Functional Anatomy of the Locomotor Apparatus. — Bewrlin: Springer-Verlag, 1980.
34. Neumann D.A. Hip abductor muscle activity as subjects with hip prostheses walk with different methods of using a cane // Phys. Tlier. — 1998. — 78. — Р. 490-501.
35. Delp S.L., Maloney W. Effects of hip center location on the moment-generating capacity of the muscles // J. Biomeeh. — 1993. — 26. — Р. 485-199.
36. Free S.A., Delp S.L. Trochanteric transfer in total hip replacement: effects on the moment arms and force-generating capacities of the hip abductors // J. Orthop. Res. — 1996. — 14. — Р. 245-250.
37. Bergmann G., Deuretzbacher G., Heller M. et al. Hip contact forces and gait patterns from routine activités // J. Biomecli. — 2001. — 34. — Р. 859-871.
38. Novacheck T.F. The biomechanics of running // Gait. Posture. — 1998. — 7. — Р. 77-95.
40. Winter D.A. Biomechanics and Motor Control of Human Movement. — 2nd ed. — New York: Wiley, 1990.
41. Paul J.P. Forces transmitted by joints in the human body // Proc. Inst. Mech. Eng. — 1967. — 181. — Р. 8-15.
42. Paul J.P. Forces transmitted by joints in the human body // Proc. R. Soc. Land. B. — 1976. — 192. — Р. 163-172.
43. Morrison J.B. Bioengineering analysis of force actions transmitted by the knee joint // Biomed. Engng. — 1968. — 3. — Р. 164-170.
44. Van den Bogert A.J., Read L., Nigg B.M. An analysis of hip joint loading during walking, running and skiing // Med. Sci. Sports Exerc. — 1999. — 31. — Р. 131-142.
45. McNitt-Gray J.I. Kinetics of the lower extremities during drop landings from three heights // J. Biomeeh. — 1993. — 26. — Р. 1037-1046.
46. Brinckmann P., Frobin W., Hierholzer E. Stress on the articular surface of the hip joint in healthy adults and persons with idiopathic osteoarthrosis of the hip joint // J. Biomeeh. — 1981. — 14. — Р. 159-156.
47. Greenwald A.S., O’Connor J.J. The transmission of load through the human hip joint // J. Biomeeh. — 1971. — 4. — Р. 507-528.
48. Rushfeldt P.D., Mann R.W., Harris W.H. Improved techniques for measuring in vitro the geometry and pressure distribution in the human acetabulum. Parts I and II // J. Biomeeh. — 1981. — 14. — Р. 253-260, 315-323.
49. Brown T.D., Shaw D.T. In vitro contract stress distributions in the natural human hip // J. Biomeeh. — 1983. — 16. — Р. 373-384.

/104-1.jpg)
/108-1.jpg)